Location: Home >> Detail
J Psychiatry Brain Sci. 2026;11(3):e260007. https://doi.org/10.20900/jpbs.20260007
 ,
Amir Kanbar 1 ,
Amish Gungah 2 ,
Bethany Waisman 3 ,
Areej Aziz 4 ,
Lexi Clark 1 ,
Megan Lopez 5,6 ,
Antonia Cheung 5,7 ,
Dingbo Wu 8 ,
Kyle Valentino 2,*
,
Amir Kanbar 1 ,
Amish Gungah 2 ,
Bethany Waisman 3 ,
Areej Aziz 4 ,
Lexi Clark 1 ,
Megan Lopez 5,6 ,
Antonia Cheung 5,7 ,
Dingbo Wu 8 ,
Kyle Valentino 2,*
1
2
3
4
5
6
7
8
*
The high prevalence and economic burden of injuries and deaths related to violence and aggression invite the need to further assess the biological mechanisms underlying their expression. Evidence implicates estrogen signaling as a mediator of aggressive behavior. In accordance with PRISMA guidelines, a systematic search was performed using the following electronic databases: PubMed, Medline, Cochrane Library, PsycInfo, Embase, Scopus, and Web of Science. The databases were searched from inception to October 3, 2025. Study screening and selection were performed by three co-authors (AK, LC, and AA). The included studies reported on the role of estrogen in aggressive and antisocial behavior. A total of 103 studies were included in this review. Preclinical and clinical studies have consistently demonstrated the role of estrogen in social dominance and antagonistic behaviors. Early-life stress and anxiety can counteract these effects. Similarly, extant data suggest that aromatase, an enzyme that synthesizes estrogen from testosterone, is involved in reported aggression. This convergence between human and animal data supports the role of estrogen in instinctive behavior and invites future studies to assess the role of estrogen antagonism in aggressive behavior.
According to the World Health Organization (WHO), 4.4 million people die from aggressive and violent acts per year [1]. In humans, anger and aggressive behavior define and are comorbid with several pathological disorders, including schizophrenia, ADHD, and conduct disorder, as well as externalizing and narcissistic behavior [2–4]. Violence-related injuries account for 10% of all years of disability [1]. The economic cost of aggression and violence in Europe and North America is estimated to be in the billions, and a 10% reduction is estimated to yield $105 billion USD in savings [1].
Existing preclinical and clinical data indicate a robust relationship between estrogen and aggression. For instance, the administration of estrogen to rodent models increases attack behavior, social dominance and territoriality [5–7]. Similarly, reducing estrogen signaling decreases aggression [8]. In humans, estrogen is associated with physical aggression, outgroup dislike and psychopathology [9,10]. When estrogen levels are high in the female menstrual cycle, females exhibit increased impulsivity [11]. From an evolutionary perspective, this is suggested to optimize maternal care and offspring defense [11]. Likewise, lactation primes female rodent defense, wherein estrogen signaling is necessary for the expression of aggressive behavior [12]. Typical male aggression is also purposive and flexible; according to human and animal data, it is typically male oriented, is territorial for the purpose of defending nutritional and reproductive resources, and requires estrogen signaling [13–15].
Epigenetic and contextual factors also play a role in the expression of aggressive behavior. For instance, aromatase, which converts testosterone into estradiol, is associated with aggression according to both preclinical and clinical data, potentially underlying the association between testosterone and male competitive and tribalistic behavior and explaining the literature suggesting that testosterone is insufficient to produce typical behaviors in males [16,17]. Estrogen may also interact with oxytocin and serotonin signaling, which have been associated with xenophobic and antisocial behavior [18,19]. Moreover, experience may modify the role of estrogen signaling in approach-avoidance behavior, where the activation of estrogen cells in the ventrolateral part of the ventromedial hypothalamus (VMHvl), the attack center of the brain, typically causes aggression; however, it causes flight following social defeat [13]. Similarly, winning previous aggressive encounters (otherwise known as the “winner effect”) is suggested to increase abnormal male aggression, including attacks on larger conspecifics, females, and vulnerable body areas, and is ablated when estrogen signaling is knocked out [20–22]. Thus, understanding the role played by nonhormonal and experiential factors in aggressive behavior will help elucidate the role of estrogen in its penetrance. Herein, the aim of this systematic review is to characterize and evaluate the effect of estrogenic signaling on aggressive behavior.
The 2020 Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines were applied in this study [23]. A systematic search was performed using the following electronic databases: PubMed, Medline, Cochrane Library, PsycInfo, Embase, Scopus, and Web of Science from inception to 2025 October 3. The search strings can be found in Table S1. The retrieved studies were independently screened by three co-authors (AK, LC, and AA) using the Covidence platform [24]. Following the removal of duplicates, the titles and abstracts of the articles were screened. Articles were required to be deemed relevant by at least two co-authors. The full texts of the articles were subsequently reviewed against the eligibility criteria (Table 1). The inclusion of articles required at least two co-authors to deem the article relevant. Discrepancies were resolved through discussion with all the coauthors.
 Table 1. Inclusion/exclusion criteria.
Table 1. Inclusion/exclusion criteria.
Published data from selected articles were extracted by A.K., L.C., and B.S. using a piloted data extraction form. The data were cross-validated with all the coauthors. Extracted data for preclinical and clinical studies are presented in Tables S2 and S3, respectively. Information to be extracted was identified a priori and included: (1) study design, (2) sample size, (3) sample age, (4) study length, (5) measurement tools, and (6) outcomes related to aggression.
Quality AssessmentTo assess the methodological quality of the included studies, risk of bias tools was applied in accordance with the study designs employed. All the studies were independently rated by at least two co-authors (AK, LC, or BS). As with the data extraction, the results were cross-validated through discussion with all the coauthors. For randomized controlled trials, the Revised Cochrane Risk of Bias tool (RoB 2) was used [25]. The Newcastle–Ottawa Scale (NOS) was used to assess cohort, case-control, and cross-sectional studies [26]. Nonrandomized studies for which the design was unclear were assessed using the cohort NOS. Preclinical in vivo studies were assessed using the SYRCLE Risk of Bias tool for Animal Studies [27].
The literature search yielded 826 studies. An additional 50 studies were identified through manual searching of reference lists. Of these, 675 studies were excluded at the title and abstract stage, including 9 duplicates, and 201 studies were assessed against the eligibility criteria outlined in Table 1. Following the assessment of full texts, 98 studies were excluded, yielding 103 included studies overall. Further details on the study selection process are depicted in Figure 1.
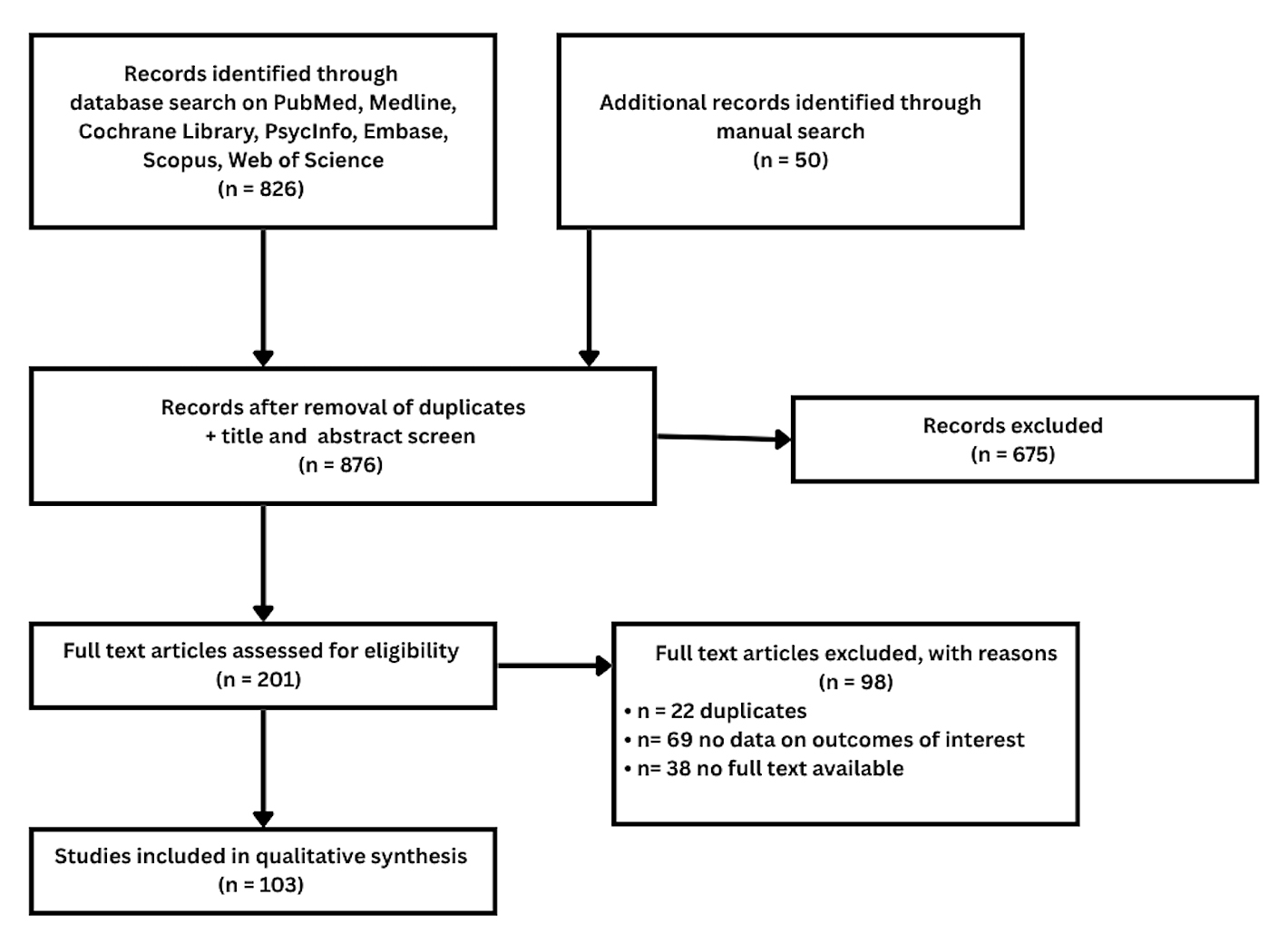 Figure 1. Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) study selection flow diagram.
Figure 1. Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) study selection flow diagram.
Among the 103 studies, 64 were preclinical. Among these studies, approximately 26 investigated estrogen signaling using genetic approaches, 15 correlated serum estrogen with aggressive behavior, and 11 assessed the effect of estrogen administration. Four studies assessed primate models, and 8 studies assessed the role of aromatase in aggressive behavior. Characteristics of the preclinical studies are summarized in Table S2.
Thirty-nine clinical studies were included in this review. Nineteen cross-sectional, 9 cohort, 5 case-control, and 6 randomized controlled trials (RCTs) were included; these studies typically assessed the correlation between aggressive and dominance-related behaviors and serum estrogen levels. Three studies assessed the role of estrogen administration. Additional studies assessed the role of genetic variants, the effect of estrogen on white matter integrity, and the role of aromatase variants in aggressive behavior. Diverse populations, including males, females, adolescents, and adult inmates, were assessed. Characteristics of the clinical studies are summarized in Table S3.
Quality AppraisalThe quality appraisal results are presented in Tables S4–S8. Animal studies typically presented some concerns, with a minority of studies presenting high risk. Scores for case-control studies varied from 6 to 8 (out of a possible maximum of 8). Awarded stars for cross-sectional studies varied from 4 to 8 (out of a possible maximum of 9). Overall, the RCTs were low risk. Cohort studies scored between 6 and 9 out of a possible 9 stars, with the majority rated as High quality. Common methodological limitations of the preclinical studies included incomplete blinding of housing conditions and outcome assessment, unclear randomization and allocation concealment, and in some cases incomplete outcome reporting. Common methodological limitations among cross-sectional studies included inadequate sample representativeness, insufficient justification of sample size, and failure to account for non-respondents. A common concern associated with the included RCTs was bias in the measurement of the reported outcome.
Preclinical Effects of Estrogen on Aggressive BehaviorAcross the majority of analyzed animal studies, the evidence suggests that estrogen signaling contributes to aggressive behavior, though findings vary across species, sex, and experimental paradigm [5,6,8,17,28–52]. Replicated studies have shown that serum estrogen levels are elevated in rodents expressing social dominance, with an inverse relationship between domesticated behavior and estrogen. For example, serum estradiol was associated with aggression in castrated males [28]. Similarly, estrogen was associated with dominant social status [7,8,29]. These effects in males were replicated in dominant females [53,54]. Replicated pharmacological findings indicate that estradiol administration increases aggressive behavior according to resident intruder (RI) paradigms [30–33]. Similarly, transcriptomic analyses have demonstrated that the expression of the estrogen receptors alpha (ERα) and beta (ERβ) is upregulated in aggressive rodents, namely, in the lateral septum, bed nucleus of the stria terminalis (BNST), and anterior hypothalamus [28,34,35]. Nonetheless, the magnitude and direction of effects vary by species, sex, and housing conditions.
Moreover, functional analyses likewise demonstrate the role of estrogen in aggressive behavior, where replicated findings indicate that inactivating estrogen signaling reduces aggressive behavior. For example, knocking out either ERα or ERβ was associated with reduced aggression in residents tested in RI paradigms [30,36]. Similarly, ERβ-KO mice failed to form stable dominance hierarchies [8]. Optogenetic activation of VMHvl estrogen (Esr1+) cells facilitated attack behavior in dominant rodents [36,37]. Similarly, optogenetic activation of BNST Esr1+ cells induced infanticide [55]. Overexpression of ERα in the medial amygdala inhibited prosocial behavior [38]. Social variables, including dominance rank, prior experience with winning or losing, and housing conditions, further modulate the expression of estrogen-dependent aggression, indicating that estrogen signaling is a contributing rather than determining factor.
Moreover, these effects of estrogen on aggressive behavior are suggested to be modified by levels of anxiety. For example, maternal separation reduced aggression in ERβ-KO mice [39]. It was also demonstrated that aversive corncob bedding can counteract the effect of estrogen on aggression and reduce the expression of ERα in the VMHvl and BNST [40]. This study demonstrates the role of development in the expression of territorial behavior. Similarly, as previously reported, the activation of VMHvl Esr1+ cells initiates attack behavior; however, activation following social defeat facilitates flight and aversive behavior [13]. Additionally, the activation of estrogen cells in the medial preoptic area (MPOA) facilitates the suppression of aggressive behavior in response to a larger conspecific [41], highlighting the role that rank and regional differences in estrogenic signaling play in the expression of adaptive aggressive behavior.
Of note, a number of preclinical studies did not observe significant effects of estrogen or estrogen receptor manipulation on aggressive behavior [36,56–60]. Possible reasons for null or suppressive findings include species- and sex-specific differences in estrogen receptor subtype distribution, variation in paradigm sensitivity, differences in housing conditions (as demonstrated by Villalon Landeros et al. (2012) [40]), and the organizational versus activational distinction in hormonal programming of aggression. Nonetheless, the convergence of correlative, pharmacological, and functional evidence across the majority of analyzed studies supports a role for estrogen signaling in aggressive behavior.
Clinical Effects of Estrogen on Aggressive BehaviorClinical studies report associations between serum estrogen and aggressive behavior, though findings vary considerably by population, sex, hormonal context, and the type of aggression assessed. For example, in university students, serum estradiol is positively correlated with aggression in men (r ≈ 0.28–0.42, p < 0.05) [14]. These effects were replicated in males reporting alcohol-related aggression [61] and in adolescent males [56]. Similarly, after controlling for age and pubertal development, free estradiol was positively associated with aggressive risk taking among adolescent girls (beta = 0.19; p < 0.01) [42]. Moreover, in menstrual cycle analyses, estradiol was positively correlated with verbal aggression (follicular phase) in healthy adult females [43]. Similarly, anger, irritability and reactive aggression significantly increased during the mid-luteal phase, with proactive aggression peaking during the follicular phase [57]. In adolescent boys, estrogen administration increased self-reported physical aggression and aggressive impulses [10]. In contrast, in small-sample dementia populations, mixed findings have emerged, where estrogen therapy was associated with a rebound in aggressive behavior in advanced male populations [62] versus a reduction in total and verbal aggression in moderate-severe female-dominated populations [63]. In contrast, other cross-sectional studies in healthy adult males found no significant correlation between estradiol and aggression measures, underscoring the moderating role of population characteristics.
Moreover, estrogen is positively associated with correlates of aggressive psychological behavior. For example, in adults, serum estradiol was significantly correlated with psychopathology in men and negativism in women [45]. Similarly, greater levels of externalizing behaviors were found to be associated with high estradiol levels; however, these behaviors were associated only under low cortisol levels and high levels of disagreeableness and emotional instability, which is consistent with the dual hormone hypothesis [46]. Similarly, using positron emission tomography, Takahashi et al. (2018) [17] reported that subcortical expression of aromatase was associated with aggressive behavior in females and self-transcendence overall in both sexes. Similarly, Górniak et al. (2025) [64] reported that grandiose narcissism was positively correlated with estradiol in women. Evidence also suggests that estrogen is positively associated with externalizing behavior and reduced empathy [46,47]. Critically, these associations are not unconditional. For instance, estradiol was associated with externalizing behaviors only in the context of low cortisol and elevated negative personality traits, consistent with the dual hormone hypothesis.
Additionally, a number of clinical studies did not observe significant positive associations between estrogen and aggressive behavior [65–68]. Reasons for null findings are multifactorial, including small sample sizes, cross-sectional designs, and heterogeneity in aggression measurement instruments. Furthermore, the relationship between estrogen and aggression appears moderated by personality traits, concurrent stress hormones, and hormonal context [46,61], which may attenuate or reverse effects in certain subgroups. Notwithstanding these findings, the balance of replicated evidence across diverse populations supports a positive association between estradiol and aggressive behavior.
The present review indicates that estrogen signaling is associated with aggressive behavior across preclinical and clinical studies; however, the strength, direction, and expression of this association are variable and depend on developmental context, sex, hormonal milieu, and experiential factors; In preclinical studies, correlative as well as pharmacological and functional analyses have indicated that estrogen signaling facilitates aggressive behavior. This effect can be counteracted by early-life stress-inducing factors such as maternal separation, cardboard bedding, and social defeat [13,40]. In humans, replicated findings indicate that estrogen facilitates aggressive and translationally relevant psychological behaviors, including narcissism and psychopathology, precursors and comorbidities to aggressive behavior [56,61,64]. Interestingly, these traits may be modified by personality and stress level [46].
Data from preclinical and clinical studies also suggest that aromatase expression is significantly associated with aggressive behavior. These findings indicate that the conversion of testosterone to estrogen facilitates male-typical behaviors typically associated with testosterone and that testosterone does not induce violent phenotypes or behavior [58,69]. However, the specific behavioral outcomes of aromatase activity appear to depend on sex and individual brain differences. These findings are consistent with functional cell type data suggesting that estrogen signaling facilitates attack postures and instinctive defense [13,30,36].
The literature indicates that diverse neuromodulator populations are associated with aggressive and antisocial behavior. For instance, oxytocin administration is associated with increased out-group derogation in humans [18]. Reward signaling has been implicated in aggression seeking and social dominance across preclinical and clinical studies [48,59]. How estrogen may synergize with these factors to produce the full repertoire of aggressive behaviors observed in humans is a vista for future research, for instance, from offspring defense to investigations related to proactive versus reactive human aggression. Additional factors that may play a role in the influence of estrogen on aggressive behavior include lifestyle factors and evolutionary life history traits. Ethnic cohorts from a fast-life-history biogeographic origin typically express ADHD, a psychiatric diagnosis that is predictive of violence and aggressive behavior [49–51,60]. In contrast, in individuals from high-latitude origins, reduced estrogen is suggested to promote the emergence of complex cognitive behavior and the expression and selection of slow-life-history traits due to reduced vitamin D-induced production of estrogen and testosterone [52,60]. This finding is consistent with hypotheses suggesting that a reduction in instinctive behaviors is associated with the emergence of modern human cultures and self-domestication [70,71].
Several limitations affecting our inferences and interpretations should be noted. First, to our knowledge, no published studies have analyzed the effect of pharmacological estrogen antagonism on aggressive behavior. Likewise, outside of 3 studies assessing estrogen administration using small sample sizes, all the included human studies were correlative. The available literature on the role of estrogen in the expression of aggression in several relevant mood disorders, including schizophrenia and ADHD, is also limited.
In conclusion, our evaluation revealed that estrogen signaling is directly and indirectly involved in aggressive behavior. Available data support an association between estrogen signaling and aggressive behavior across human and animal studies, though this relationship is moderated by contextual and individual-level factors, including hormonal interactions, social experience, and personality traits. Although the full mechanistic model of estrogen’s effects is unknown, the conversion of testosterone into estradiol is implicated and should be included in discussions regarding the influence of sex hormones on aggressive behavior. Likewise, future studies should assess the role of estrogen antagonism on aggression, including analyses on neuromodulator populations implicated in aggression (e.g., oxytocin and dopamine). Further studies should also investigate the correlation between the winner effect, estrogen and aggression in human populations. Likewise, preclinical aggression-related suicidality models may benefit from investigations into estrogen signaling.
Not applicable. This systematic review analyzed previously published studies and did not involve direct human or animal subjects.
Declaration of Helsinki STROBE Reporting GuidelineNot applicable. This study is a systematic review and did not involve direct human subject’s research.
The following supplementary materials are available online. Table S1: Search Terms; Table S2: Summary of characteristics of the preclinical studies; Table S3: Summary of characteristics of the clinical studies; Table S4: Results of study quality appraisal of included randomized controlled trials; Table S5: Results of study quality appraisal of included cohort studies; Table S6: Results of study quality appraisal of included cross-sectional studies; Table S7: Results of study quality appraisal of included case-control studies; Table S8: Results of study quality appraisal of included animal studies; Table S-Ref: Complete reference list of all 103 analyzed studies.
All data analyzed in this study are available within the manuscript and supplementary materials.
Conceptualization, KV; Methodology, KV and BMO; Investigation/Data Curation, AK, LC, AA, BW, BMO, AG, ML, AC, DW and KV; Writing Original Draft, KV, AK; Writing Review & Editing, BMO and KV; Supervision, KV.
The authors declare no conflicts of interest.
This research received no external funding.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
Oană BM, Kanbar A, Gungah A, Waisman B, Aziz A, Clark L, et al. The Role of Estrogenic Mechanisms in Pathological Aggression: A Systematic Review of Preclinical and Clinical Trials. J Psychiatry Brain Sci. 2026;11(3):e260007. https://doi.org/10.20900/jpbs.20260007.

Copyright © Hapres Co., Ltd. Privacy Policy | Terms and Conditions




